 病原菌抗药性机制:病原真菌的抗药性是病原真菌力图躲避、抵制和消除杀菌剂对其作用而获得的适应性代谢变化。这种变化归纳起来有其遗传和生化两大机制。
病原菌抗药性机制:病原真菌的抗药性是病原真菌力图躲避、抵制和消除杀菌剂对其作用而获得的适应性代谢变化。这种变化归纳起来有其遗传和生化两大机制。
1. 遗传机制 病原体抗药性性状是由遗传基因决定的。抗药性基因可能存在于细胞核中的染色体上,也可能存在于细胞质中(例如线粒体上)。已知绝大多数抗药性基因位于细胞核的染色体上。
(1) 主效基因抗药性 植物病原体对某种杀菌剂的抗药性是由一个主基因控制的称为单基因抗药性,或主效基因抗药性。已知目前病原菌对杀菌剂的抗药性大多数都属于单基因抗药性。该基因可能是一段由若干核苷酸组成的DNA片段,其中单个或几个核苷酸的改变均能通过相同的生化机制表达对药剂的抗药性,但是不同的核苷酸改变可能表达不同的抗药性水平和适合度,这就是等位基因抗药性。一种植物病原体群体中可能同时存在着多等位基因抗药性。
(2) 寡基因抗药性 植物病原体细胞中可能存在几个主效基因可以决定对一种药剂的抗药性,其中任何一个基因发生突变即可表达抗药性。植物病原体同一个体可能发生一至几个这种主效基因的变异,不过其中一个突变基因对另一个突变基因往往具有上位显性作用。尽管它们之间可能发生相互作用而表现型不同于单基因突变体,但是抗药性水平通常与单基因抗药性表达的抗药性水平相似,这就是寡基因抗药性。
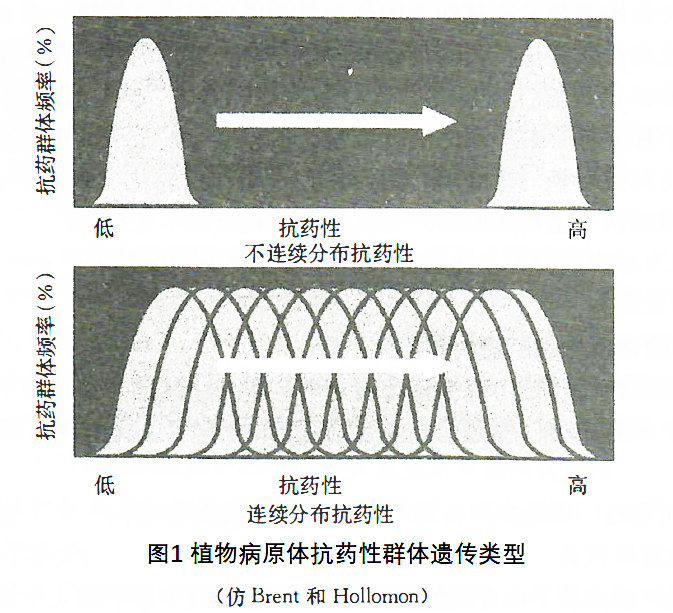
与敏感病原体等位基因相比,每个突变基因可能表现为完全显性或不完全显性,或完全隐性或不完全隐性。当同一病原体个体细胞中存在等位的敏感基因和抗药性基因时,其表现可能是敏感的或抗药性的。例如卵菌及其他双倍体阶段致病的植物病原体,只有当控制抗药性的基因是显性的,或隐性基因的纯合体才能表达抗药性。大多数植物病原体子囊菌、担子菌和半知菌的致病阶段是单倍体阶段,决定抗药性的基因无论是显性、半显性,还是隐性均能表达抗药性。主基因或寡基因控制的抗药性,抗药性水平往往很高,抗药性和敏感个体杂交后代对药剂的敏感性表现为抗药性和敏感不连续的孟德尔遗传分离规律(图1)。使植物病原体表现质量性状的抗药性基因往往是药剂的靶标基因,在生产上出现质量性状抗性的杀菌剂有苯并咪唑类、苯酰胺类、羧酰替苯胺类、苯胺基嘧啶类、甲氧基丙烯酸酯类、二甲酰亚胺类等杀菌剂和春雷霉素、链霉素等抗生素。
(3) 聚基因抗药性 有些植物病原体对少数药剂的抗药性是由许多微效基因的突变引起的。这些微效基因的作用可以相互累加,使抗药性水平显著增加,这就是聚基因 (polygene) 抗药性。抗药性与敏感菌株的杂交后代中不同基因型组别重叠,对药剂的抗药性水平差异是连续的,表现为数量遗传。即使在药剂的长期选择压力下,植物病原体群体敏感性仍然保持连续分布,只是整个分布向降低敏感性和增加抗药性水平的方向数量移动(图1)。使植物病原体表现数量遗传性状的抗药性基因往往是非靶标基因,在生产上出现数量性状抗性的杀菌剂有多果定、放线菌酮、三唑醇、三唑酮、咪鲜胺、氯苯嘧啶醇等麦角甾醇生物合成抑制剂。
(4) 多重抗药性 植物病原体可以同时或先后对不同类型的农药产生不同的抗药性基因突变而表现多重抗药性。其中各个基因的突变及其调控的生化机制是独立的,互不干扰,这就是多基因 (multigene) 抗药性。例如灰葡萄孢霉 (Botrytis
cinerea) 可以对苯并咪唑类、二甲酰亚胺类、苯胺基嘧啶类及乙霉威等不同类型的杀菌剂产生多重抗药性。表现多重抗药性状的植物病原体往往是先后或同时发生了不同类型杀菌剂的靶标基因抗药性变异。
(5) 胞质基因抗药性 植物病原体抗药性基因还可能存在于细胞质中的线粒体、质粒、整合子或病毒分子上。在实验室通过菌体线粒体DNA的突变,可以获得对氯霉素、放线菌酮、链霉素等抗生素的抗药性。但是实际情况下,抗生素的抗药性基因似乎很少位于线粒体或染色体上。
丝状病原真菌的多核菌丝可能是异核的,即同一细胞内的细胞核不具有遗传同一性。异核体的不同细胞核可能包含对某种杀菌剂抗药性和敏感的等位基因。它们能表达各自控制的性状,在有药或无药条件下均能正常生长,但随着药剂选择压的变化,菌体内抗药性和敏感的细胞核比例可能发生改变。
(6) 抗药性基因的多效作用 植物病原体抗药性基因往往具有多效作用。即基因发生抗药性突变,也可能同时引起其他表现型特征的变化,这是基因之间特异性互作发生改变的结果。例如控制二甲基亚胺类杀菌剂抗药性的高抗基因,通常会引起菌体对高渗透压的超敏感性,三唑醇抗药性基因还会引起菌体生长减慢、产孢减少等。(引自植物化学保护学,略有改动)
2. 生化机制 主要表现:
(1) 病原真菌细胞产生某种变化。如降低原生质膜的透性,使杀菌剂不能达到作用位点,例如灭瘟素和多抗霉素的作用机制是通过抑制几丁质合成酶而干扰几丁质的合成。在抗性菌株提取液中的这两种药剂,仍能抑制几丁质的合成,因此其抗药性可能是原生质膜透性降低,药剂达不到作用点所致。
(2) 病原真菌增强了对杀菌剂的解毒能力,使杀菌剂降低或丧失活性。对五氯硝基苯产生抗药性的镰刀菌,能把五氯硝基苯转化成低活性的五氯苯胺和五氯硫酚即是一例。
(3) 病原菌通过代谢变化,阻止了杀菌剂在体内的活化作用。吡菌磷的杀菌作用是通过病原菌将其转化成杀死病原真菌的2-羟基-5-甲基-乙氧羰-吡唑并(1,5-a)嘧啶,而抗性菌则通过代谢的改变,降低了这种转换作用的能力。
(4) 亲和力下降。杀菌剂虽然能到达作用位点,但降低了作用位点对杀菌剂的亲和力。苯菌灵、多菌灵、甲基硫菌灵和放线菌酮的抗药机制均属这类。其作用点上的微小变化,均能降低对杀菌剂的亲和力,而产生抗药性。
(5) 迂绕作用。病原真菌通过改变代谢,从旁路绕过杀菌剂阻碍的代谢反应,使其不能发挥杀菌作用。
(6) 补偿作用。某些杀菌剂以特异性酶作为主要作用位点时,病原真菌通过增加酶的数量,以弥补因药剂作用的损失,使整体代谢正常。